Global Review of Social Indicators used in Protected Area Management Evaluation
By Josh Ratay, SRC Intern
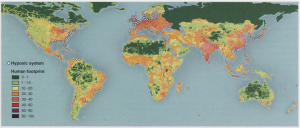
Global map of the world’s protected areas. These areas are numerous and often require a great deal of management assessment, in which social indicators play a large role. Image from University of Maryland, Baltimore County, using data from the United Nations Environment Programme.
Although conservation deals heavily with environmental data, social aspects of conservation initiatives are also extremely important. When human-related considerations are taken into account, certain trade-offs must often be made, but the end result is usually more effective at fulfilling the goals of the project, particularly at the local level. In a recent study, researchers examined the social aspects of protected areas around the world, relating social indicators and well-being dimensions. These went beyond simple measures of wealth and included education, healthcare, government, and social unity.
A pre-existing database, the Global Protected Area Management Effectiveness Database (GDPAME), which contains records of assessments made at different protected areas around the world. It contains information about both social and environmental factors and is used by policymakers as a source of information. For the study, social indicators were any factors that were placed in the context of local/indigenous affairs. Multiple existing frameworks were used to analyze indices of well-being. Indicators were examined in different scenarios, and classified as positive, negative or neutral. Differing methodologies for assessing protected areas very greatly in the number of indicators used, ranging from 13 to 429. The percentage of indicators relating to social issues also vary greatly, with a minimum of 7% and a maximum of 60%. Interestingly, the methodology which used 60% social indicators was exclusively for marine protected areas (“How is Your MPA Doing?”).

“How is Your MPA Doing?”, a manual released as a collaboration between NOAA, WWF, the IUCN and others, serves as a method for evaluating MPAs which focuses heavily on social indicators.
The study found that the most common dimension of well-being to be taken into account was living standards. Security and culture were also high-ranking. Overall, about one quarter of all indicators were social, showing that conservation management is increasingly taking social considerations into account. However, the study did recommend additional actions that should be taken to further increase coverage. Firstly, it proposed greater focus on community dynamics, as opposed to purely physical measurements such as resource availability. Additionally, many of the indicators focused heavily on the state of a conservation project after its implementation, and put less emphasis on local engagement during the early phases of an initiative. Changing this could better promote community participation by involving locals in the building of a policy, instead of only the continued management of existing policies. Additionally, the cultural values of a given area should be given increased consideration. The study also noted the bias toward describing environmental impacts of human initiatives as negative, and recommended that more specific language be used for future indicators. Overall, this review of social indicators and well-being dimensions showed the importance of integrating local communities in both the planning and assessment methods of protected areas to improve the effectiveness of conservation initiatives.
Work Cited
Corrigan, Colleen, et al. 2016. “Global Review of Social Indicators used in Protected Area Management Evaluation.” Conservation Letters.