Permeable Pavement Systems as a Mitigation Strategy to Combat Stormwater Outfall and Sea Level Rise
By Casey Dresbach, SRC intern
The EPA defines stormwater runoff as that which is generated from rain events that flow over land or impervious surfaces, such as paved streets, parking lots, and building rooftops, and does not soak into the ground (EPA, 2018). This accumulation of water ultimately ends up in coastal waters, but upon its journey it picks up a plethora of pollutants ranging from trash, chemicals, fecal matter, and sediment. These substances picked up by the runoff are compromising the health and water quality of these waters. A large portion of the global warming heat is accumulating in oceans worldwide. As a result, sea levels are rising through thermal expansion of water in the oceans and melting of ice sheets and glaciers on land.
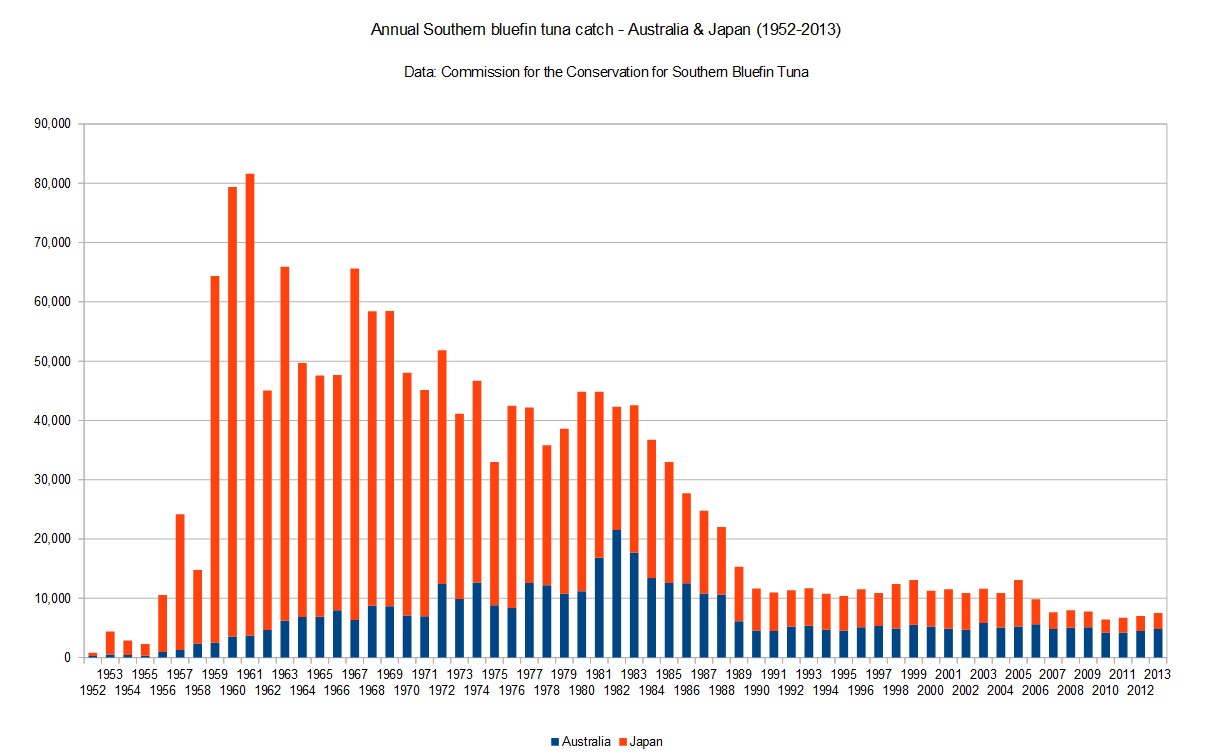
Between 1998 and 2006 alone, tidal gauges showed sea levels in South Florida followed average global rate (0.04 to 0.20 inches per year) – but in 2006, local rates suddenly underwent a rapid acceleration, averaging about 0.20 inches to a half inch each year (Staletovich, 2016). Over the last decade, flood management has become increasingly important to several coastal states at risk such as Florida, Louisiana, New York, and California with each region varying in susceptibility to sea level rise. Flood management is commonly issued by drainage systems such as complex pipe networking. However, these projects are extremely costly and only seem to be dealing with the issue on a short-term basis. These stormwater controls are known as best management practices (BMPs) intended to filter out pollutants. Miami Beach has already undergone a major BMP using “gray stormwater infrastructure,” that is their $500 million drainage and water treatment system, one of which resides in a highly populated area (Harris, 2018). This coastal area, also known as Sunset Harbor Village, is home to local businesses, restaurants, bars, and boutique fitness enterprises. Yet, the effectiveness is not justifying the efforts taken to mitigate the impacts of sea level rise and urban development. The pump fails to filter out many of these pollutants pervading the streets and they ultimately end up into the coastal waters of Biscayne Bay (Figure 1).

Figure 1. Dirty Water Pumped Into Biscayne Bay (Caption: Dirty water pumped into Biscayne Bay in Miami Beach, FL. Snapshot from video taken by Miami Beach citizen. (Lipscomb, 2019))
Benjamin O. Brattebo and Derek B. Booth conducted a study to examine the long-term stormwater quantity and quality performance of permeable pavement systems. Permeable pavement is made up of a matrix of concrete blocks or plastic structure with holes filled with sand, gravel or soil (Brattebo & Booth, 2003). The filling absorbs the outfall that might otherwise percolate into alternative places, most notably are coastal environments. The voids allow stormwater to seep through the pavement and into the soil. Asphalt pavement, an impervious system, was compared to four permeable pavements created in a monitored and controlled parking lot in Renton, Washington. Employees of King County Public Works facility utilize the parking facility during the workweek, Monday through Friday. Plastic Grasspave, plastic Gravelpace, concrete Turfstone, and concrete UNI Eco-stone were the permeable systems used. The first two plastic grids differed in their fill. The first was filled with sand and planted with grass and the second was filled with gravel only. Similarly, the first concrete was filled with soil and planted with grass while the latter was filled with gravel. Each permeable pavement had two parking stalls paired into one instrument station (Brattebo & Booth, 2003), which made up 8 stalls. The ninth stall was the asphalt-paved control. See Figure 2. During 15-minute rainfall events, runoff rates were cataloged. Water was collected and sampled in both the impermeable and permeable systems. Surface durability, infiltration capacity, and water-quality performance were assessed. Results showed that overall samples from permeable systems contained overwhelmingly lower levels of copper and zinc, and none had traces of diesel fuel or lead. Researchers indicated that while their findings support long-term success for infiltration via permeable systems, the success might not be universal in places with extremely arid or colder snow induced climates.
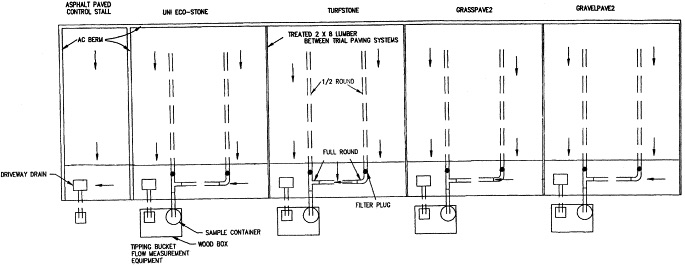
Figure 2. Plan View of 9 Parking Stalls JPEG aligned center. (Caption: 8 impermeable systems installed in parking lot alongside 1 control, the permeable asphalt concrete system. (Brattebo & Booth, 2003))
More research needs to be done in order to combat the challenges that arise with stormwater runoff and urban redevelopment continues on an upward trend. Long-term solutions and studies are crucial but as are short-term. A perfect balance of the two will help mitigate the detrimental impacts of pollutants trickling into larger water bodies that are utilized by the overall marine ecosystem, humans and coastal fauna included.
Works Cited
Brattebo, B. O., & Booth, D. B. (2003). Long-term stormwater quantity and quality performance of permeable pavement systems. Water Research , 37 (18), 4369-4376.
EPA. (2018, September 14). NPDES Stormwater Program. Retrieved from EPA: https://www.epa.gov/npdes/npdes-stormwater-program
Harris, A. (2018, April 18). Miami Beach’s future is ‘uncertain,’ experts say, but sea rise pumps are a good start. (Miami Herald) Retrieved March 20, 2019, from Miami Herald: Miami Beach’s future is ‘uncertain,’ experts say, but sea rise pumps are a good start: http://www.miamiherald.com/news/local/community/miami-dade/miami-beach/article209328849.html#storylink=cpy
Lipscomb, J. (2019). Videos Show Dirty Stormwater Pumped Into Biscayne Bay and Swallowed by Manatee. (M. N. Times, Producer) Retrieved from Miami New Times: https://www.miaminewtimes.com/news/videos-show-dirty-stormwater-pumped-into-biscayne-bay-and-swallowed-by-manatee-11068892
Staletovich, J. (2016, April 7). Miami Beach flooding spiked over last decade, UM study finds. (Miami Herald) Retrieved April 30, 2018, from Miami Herald: http://www.miamiherald.com/news/local/environment/article70145652.html








{kind=link}