By: Hannah Calich, RJD Graduate Student
For many years we lived in a world where the state of our fish stocks was not a primary concern. However, our population has become so large, and our technology so advanced, that we are utilizing resources at a rate that was once inconceivable. We have significantly impacted many of the world’s fish stocks and it is no longer biologically or economically feasible to continue harvesting them at this rate. Thus, we must develop ways to relieve pressure on wild fish stocks while continuing to provide the world with the fish protein it requires. Given the projected human population, and the current status of wild fish stocks, it will be up to the aquaculture industry to help the ocean meet the world’s demand for fish protein.
The global human population has been projected to reach over 9 billion people by 2050, and over 10 billion people by 2100 (Figure 1; UN, 2012). While the rate of population growth has been slowing, and may eventually reach a plateau, feeding a population of 9 to 10 billion people represents a significant challenge.
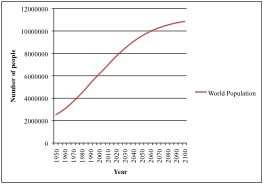
Projected world population based on a medium population growth variant (data from UN, 2012)
Along with the world’s population, meat consumption has been steadily rising. In the 1950s the world’s population was consuming 44 million tonnes annually; by 2009 that figure had increased to 272 million tonnes. When the increase in human population is taken into consideration those figures suggest that the annual per capita meat consumption rate has more than doubled, to almost 90 pounds per person (Brown, 2011).
While on average the world’s meat consumption has been rising, there are regional trends in who can afford to eat meat. As Brown (2011) said, “wherever incomes rise, so does meat consumption” (pg. 173). As a result of incomplete or insufficient diets, currently 800 million people suffer from chronic malnourishment worldwide (FAO, 2014).
Fish are an important source of protein, nutrients and energy, particularly in poorer nations where essential nutrients are often scarce (FAO, 2014). For example, 150 g of fish protein can provide 50-60% of an adult’s daily protein requirement (FAO, 2014). In addition to being nutritious, fish are often an affordable source of protein. As such, nearly 17% of the global population’s protein intake comes from fish, though again the trends are regional and that number is closer to 50% for some developing countries in Africa and Asia (FAO, 2014). Following the trend of the world’s meat consumption, per capita fish consumption as also increased, from 10 kg in the 1960s to over 19 kg in 2012 (FAO, 2014). Per capita fish consumption has increased in both developing regions (5.2 kg in 1962 to 17.8 kg in 2010) and low-income food-deficit areas (4.9 kg to 10.9 kg) (FAO, 2014). While developed nations still consume more fish, the gap is narrowing.
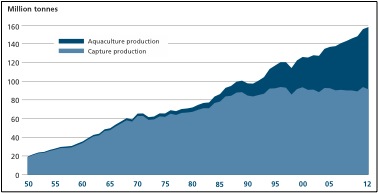
The surge in fish consumption has left the marine capture industry struggling to meet the world’s demand for fish protein. Global marine capture fisheries have been consistently harvesting between 80 and 90 million tonnes per year since the mid-1980s (Figure 2; FAO, 2014; Pauly & Froese, 2012). However, this stability is not due to stable fish populations. Instead, the stability is due to pushing the boundaries of the ocean’s fish stocks. Specifically, the marine capture industry has been targeting less desirable species, fishing further offshore, and harvesting smaller fish than ever before.
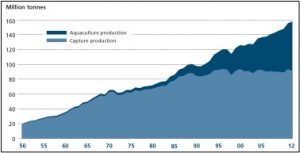
Aquaculture and capture fisheries production in millions of tonnes from 1950-2012 (Source: FAO, 2014)
This high demand for fish protein has put a significant strain on wild fish populations. Currently, approximately 30% of wild stocks are considered overfished, 60% are fished at (or close to) their maximum sustainable limit, and only 10% are being fished under their limit (FAO, 2014). Overfishing not only negatively impacts the ecosystem, but also reduces fish production and has negative social and economic consequences. For example, in areas of poor governance fishers that are unable to legally catch their quotas occasionally turn to illegal, unregulated or unreported fishing techniques to earn a living. These practices can be wasteful, dangerous and negatively impact communities and the environment (FAO, 2014). Simply put, the world’s oceans cannot continue to support the planet’s increasing demand for fish.
Aquaculture has the potential to be the solution to the world’s fish shortage. Global aquaculture production is one of the fastest growing food-producing sectors. In 2012 aquaculture provided almost 50% of all fish for human consumption and has been predicted to provide 62% by 2030 (Figure 2; FAO, 2014). Not only are we raising more fish, but we are also eating more of the fish we raise. The amount of fisheries production used by humans for food has increased from about 70% in the 1980s to move than 85% (136 million tonnes) in 2012 (FAO, 2014). In fact, in 2012 aquaculture production was higher than beef production (66 million tonnes compared to 63 million tonnes) (Larsen & Roney, 2013). This increase in productivity is largely due to an increase in small-scale fish farms (FAO, 2014).
In addition to helping feed the world, aquaculture can play a critical role in the economy. Together, the fisheries and aquaculture industries help support the livelihoods of 10-12% of the world’s population (FAO, 2014). Additionally, fish is one the most commonly traded commodities worldwide and was worth almost 130 billion dollars in 2012; a value that has been predicted to increase into the future. The fish trade is particularly important in developing nations where in some cases the trade is worth over half of the total value of traded commodities. Aquaculture also benefits to the economy because of how efficiently herbivorous fish convert feed into live weight. For comparison, the grain to live weight ratio of cattle is approximately 7:1, it is 4:1 for pork, 2:1 for chicken and less than 2:1 for herbivorous fish such as tilapia or catfish (Brown, 2006). Since herbivorous fish have such a low ratio, focusing on them will allow us to harvest more protein while using less grain.
Aquaculture, in addition to directly providing food, can also serve as a way to support and replenish natural fish stocks. For example, in 2011 the Mississippi Department of Natural Resources raised and released 7,500 cobia in the northern Gulf of Mexico. The project’s aim was to help replenish stocks and to examine the feasibility of releasing fish that were raised in aquaculture facilities into the wild as part of a stock enhancement program (Mississippi Department of Natural Resources, 2011).
While aquaculture has shown great promise, like any industry it has it’s flaws. Two of the primary environmental concerns with aquaculture are preventing ecosystem degradation, and raising carnivorous fish without harming prey populations. Ecosystem degradation comes in many forms but can include: habitat destruction, disease, pollution, and changes to a species’ population genetics. Raising carnivorous fish, such as salmon or tuna, is controversial because while they are economically important species, their growth depends on the availability of large quantities of small prey fish, such as pilchards. Harvesting large quantities of wild prey fish for fish meal can seriously impact the prey’s natural populations (Brown, 2006). To continue to grow sustainably the aquaculture industry needs to reduce its environmental impact as well as become less dependent on wild fish for feed, and increase the diversity of culture species.
While there are important sustainability concerns surrounding the aquaculture industry, the industry is progressing and adapting at a very fast rate and fortunately, these concerns are becoming less relevant. Since the world’s population is only predicted to increase, finding a way to meet the world’s demand for fish without relying on wild stocks is essential. So long as the aquaculture industry continues to develop in a way that is environmentally sustainable, aquaculture will have an important role to play in providing healthy fish protein and jobs for the world’s economy.
References:
Brown, L. R. (2006). Plan B 2.0: Rescuing a Planet Under Stress and a Civilization in Trouble. Washington, DC: Earth Policy Institute. Retrieved September 4, 2014 from http://www.earth-policy.org/books/pb2
Brown, L. R. (2011). World on The Edge. Washington, DC: Earth Policy Institute. Retrieved September 4, 2014 from http://www.earth-policy.org/images/uploads/
book_files/wotebook.pdf
FAO. (2014). The State of World Fisheries and Aquaculture 2014. Retrieved September 4, 2014 from http://www.fao.org/3/a-i3720e.pdf
Larsen, J., & Roney, M. (2013). Farmed Fish Production Overtakes Beef. Retrieved September 4, 2014 from http://www.earth-policy.org/plan_b_updates/2013/ update114
Pauly, D., & Froese, R. (2012). Comments on FAO’s State of Fisheries and Aquaculture, or ‘SOFIA 2010’. Marine Policy, 36(3), 746-752.
Mississippi Department of Marine Resources. (2011). Cobia Released on Popular Mississippi Offshore Artificial Reef. Retrieved September 4, 2014 from http://www.dmr.state.ms.us/index.php/news-a-events/recent-news/272-11-131-lst
UN. (2012). World Population Prospects: The 2012 Revision. Retrieved September 4, 2014 from http://esa.un.org/unpd/wpp/unpp/panel_population.htm